
被窃或疑病妄想简介:阿尔茨海默病性痴呆可有片断地迫害妄想、被窃或疑病妄想。阿尔茨海默病性痴呆(dementia in Alzheimer’s disease)是一种起病隐袭的进行性发展的神经系统退行性疾病。临床上以记忆障碍、失语、失用、失认、视空间技能损害、执行功能障碍以及人格和行为改变等全面性痴呆表现为特征,病因迄今未明。 过去将65岁以前发病者,称早老性痴呆(presenile dementia);65岁以后发病者称老年性痴呆(senile dementia)。本病病理改变主要特征为大脑皮质萎缩、神经原纤维化和脑神经细胞变性及老年斑,是老年期较常见的疾病。
症状起因:
(一)发病原因
从现有的流行病学资料来看,AD可能是一组异质性疾病,在多种因素(包括生物和社会心理因素)的作用下才发病。虽然AD神经病理学,特别是分子生物学研究有了很大进展,为AD病理生理和病因学研究奠定了基础,但仍处于探索阶段,AD的病因尚远未阐明。流行病学研究分析AD的危险因素,为寻找病因提供了线索,但危险因素并非病因。从目前研究来看,AD的可能因素和假说多达30余种,如家族史、女性、头部外伤、低教育水平、甲状腺病、母育龄过高或过低、病毒感染等。都是同一个问题不同侧面的探讨,可能都是正确的,并不互相排斥。从目前研究来看,AD可能存在不同原因,下列因素与本病发病有关:
1.家族史 绝大部分的流行病学研究都提示,家族史是AD的危险因素。某些患者的家属成员中患同样疾病者高于一般人群,此外还发现先天愚型患病危险性增加。进一步的遗传学研究证实,本病可能是常染色体显性基因所致。最近通过基因定位研究,发现脑内淀粉样蛋白的病理基因位于第21对染色体。可见痴呆与遗传有关是比较肯定的,但遗传的作用到底有多大还很难肯定。由于AD的发病年龄较晚,目前还没有基于普通人群的孪生子研究报道。少数老年单卵孪生子(MT)研究报道的同病率并不是很高。多数报道提示AD存在家族聚集现象,AD与一级亲属阳性家族史的关系也比较肯定。据现有资料,8篇病例对照研究,AD与一级亲属痴呆史关联显著,另一篇未发现二者关联。对欧洲的11项病例对照研究的再分析显示,如果至少有一位一级亲属患痴呆的话,则痴呆的患病危险增加3倍以上。对载脂蛋白E(Apo E)基因型在人群中分布频率的研究,进一步支持遗传因素对AD的发病作用。已经证明Apo E等位基因ε4是AD的重要危险因素。Apo E ε4基因的频率在家族性和散发性AD中都明显增高。Apo E ε4基因在尸解证实的AD患者中的频率为40%左右,而在正常对照人群中约为16%,带一个ε4等位基因患AD的危险是普通人群的2~3倍,而携2个ε4等位基因的患病危险约为普通人群的8倍。现在已经清楚Apo Eε4等位基因并不是AD发病的必备因素,它对AD发病的预测作用还有待前瞻性研究来证实。该作者分析可能与收集的病例都是晚发AD之故,提示家族聚集可能是早发AD的一个重要危险因素。但这种对阳性结果的解释宜慎重,家族聚集并非真正的遗传因素。由此可见,遗传因素并不是AD发病的惟一因素。
2.一些躯体疾病 如甲状腺疾病、免疫系统疾病、癫痫、偏头痛等,曾被作为AD的危险因素研究。有甲状腺功能减退史者,患AD的相对危险度为2.3。AD发病前有癫痫发作史较多(相对危险度为1.6)。偏头痛或严重头痛史与AD无关。不少研究发现抑郁症史,特别是老年期抑郁症史是AD的危险因素。最近的一项病例对照研究认为,除抑郁症外其他功能性精神障碍如精神分裂症和偏执性精神病也有关。比较早年中枢神经系统感染史,例如脑炎、脑膜炎、疱疹病毒感染,以及家畜接触史,食用动物脑史等未能证明这些因素与AD有关。曾经作为AD危险因素研究的化学物质有重金属盐、有机溶剂、杀虫剂、药品等。铝的作用一直令人关注,因为动物实验显示铝盐对学习和记忆有影响;流行病学研究提示痴呆的患病率与饮水中铝的含量有关。Michel等根据法国一项痴呆患病率研究的初步结果报道,铝是AD的患病危险因素,但进一步分析后又否认了此结果。Flaten等(1990)报道饮水中铝的含量与痴呆相关。此后有多项研究未能证实铝是AD的危险因素。对重金属接触史包括接触铝的人进行病例对照研究也没有发现哪一种重金属与AD有关。可能由于铝或硅等神经毒素在体内的蓄积,加速了衰老过程。但铝虽然是一种神经毒性物质,但就已有的研究看,它还不能作为AD的患病危险因素。有报道发现吸烟非但不是AD的危险因素,相反对AD起保护作用。而有作者未发现二者关联。烟民患AD少可能由于他们平均寿命短,未到老年即夭折之故。
3.教育水平 低教育水平与痴呆的患病率增高有关的报道越来越多。上海报道痴呆和AD的患病率文盲为6.9%,学龄大于6年为1.2%。最近意大利一篇流行病学调查也有类似发现。但一篇日本病例对照研究,未发现教育水平与痴呆和(或)痴呆亚型之间关联。对此尚无合理解释,有些学者认为这是因系统误差造成的。由于多数流行病学研究都是采用二阶段筛查检查法,对筛查阳性的患者再进行诊断性检查,这样文盲或文化程度低的人可能在筛查阶段认知测验得分低,容易进入诊断性检查阶段并诊断为痴呆,使得患病率增高,而事实上这些人可能根本就没有认知功能下降。认为这是由于文盲本身生物学特征所决定,而并非教育问题,而且教育与社会经济状况有关,进一步使这个问题复杂化了。然而,张明园等(1990)在筛查时根据筛查对象的文化程度不同,采用不同的筛查分界值,避免了这种系统误差,结果低教育水平者痴呆的患病率仍较高。此后有数项研究证实了这一结果。低教育水平与AD的病因联系仍不太清楚,可能的解释是早年的教育训练促进了皮质突触的发育,使突触数量增加和“脑贮备”(brain reserve)增加,因而推迟了痴呆的诊断时间。这一假说得到了一些临床观察的支持,例如,高教育水平的AD患者,即使在晚期仍可保留一些认知功能,他们从确定诊断到病死的病程相对较短。低教育水平与血管性痴呆及其他继发性痴呆也有相似的关系。
4.头部外伤 头部外伤指伴有意识障碍的头部外伤,脑外伤作为AD危险因素已有较多报道。12篇病例对照研究,有3篇发现显著关联;4篇AD既往外伤史较对照组多,但无统计学显著性,其余5篇未发现二者有任何关联。但最近一项严重脑外伤的随访研究报道,更加引起了人们的兴趣。Robert等平均随访严重脑外伤患者25年,结果有大约1/3的患者脑组织中出现类似于AD的β淀粉样蛋白沉积。临床和流行病学研究提示严重脑外伤可能是某些AD的病因之一。就目前资料来看,头部外伤可能是AD的一个危险因素,但还不能肯定。
5.母育龄过高或过低(大于40岁或小于20岁) 由于先天愚型(Down’s syndrome,DS)可能为AD危险因素,而DS危险随母育龄增高而上升。有9篇病例对照研究,有的发现有关联,有的发现有差异但无统计学显著性,或根本未发现二者相关,有的则认为仅是某些散发型AD的危险因素。
6.其他 免疫系统的进行性衰竭、机体解毒功能削弱及慢病毒感染等,以及丧偶、独居、经济困难、生活颠簸等社会心理因素可成为发病诱因。
(二)发病机制
1.遗传因素 DS有AD类似病理改变,DS如活到成人发生AD几率约为100%,已知DS致病基因位于21号染色体,乃引起对AD遗传学研究极大兴趣。但AD遗传学研究难度大,多数研究者发现患者家庭成员患AD危险率比一般人群约高3~4倍。St.George-Hyslop等(1989)复习了AD家系研究资料,发现家庭成员患AD的危险,父母为14.4%;同胞为3.8%~13.9%。用寿命统计分析,FAD 一级亲属患AD的危险率高达50%,而对照组仅10%,这些资料支持部分发病早的FAD,是一组与年龄相关的显性常染色体显性遗传;文献有一篇仅女性患病家系,因甚罕见可排除X-连锁遗传,而多数散发病例可能是遗传易感性和环境因素相互作用的结果。
双生子研究是临床遗传学研究重要方法,Kallman(1956)研究了108对双生子同病率,单卵双生为42.8%,双卵双生为8%。由于某些单卵和双卵双生子发病年龄差距甚大,不同病双生子以后可能同病。因此,单卵双生和双卵双生同病率估计比较接近约为40%,即双生子之一患AD,另一同胞患AD的几率大约为40%,但该同胞发病时间可能要晚得多。
分子遗传学技术的发展为AD的病因学研究提供了广阔前景。与AD有关的遗传学位点,目前已知的至少有以下4个:早发型AD基因座分别位于2l号染色体、14号染色体和1号染色体。相应的可能致病基因为APP、S182和STM-2基因。迟发型AD基因座位于19号染色体,可能致病基因为载脂蛋白E(Apo E)基因。提示AD遗传的异质性,即不同的DNA改变引致同一临床表现,或基因型不同而表型相同,分述如下:
(1)早发型AD:多数新资料来自少见的早发型,具有AD家族史或DS病人的研究。在与AD有关疾病基因的研究中,已取得了相当可观的进展,已发现3个基因的突变与早发型家族性常染色体显性遗传的AD有关。21号染色体的APP基因突变与β淀粉样蛋白序列相邻。APP是位于细胞膜、高尔基复合体和囊泡上的跨膜蛋白,β淀粉样蛋白部分位于膜内,部分位于膜外。在细胞培养和转基因鼠中,突变的APP表达导致β淀粉样蛋白产生增多,SAPP产生减少。β淀粉样蛋白升高会促进老年斑的形成,并与其毒性有关。APP突变引起的神经元蜕变后果中,SAPP水平下降可能扮演重要角色,因为SAPP能促进神经元的生存和增加其可塑性,还能保护神经元不受兴奋毒性物质和氧化损伤。大部分的早发型家族性AD并不是由APP突变引起。另外两个与早发型家族性AD有关的是早老素1和早老素2基因(PS1、PS2)。PS-1位于14号染色体,PS-2位于1号染色体。PS-1和PS-2表达也是跨膜蛋白,它们位于大脑神经元的内质网上。表达早老素的细胞培养和转基因鼠研究发现,早老素突变能增加β淀粉样蛋白的产生量,并增加神经元对代谢和氧化伤害的敏感性。早老素基因突变引起病理活动可能包括扰乱内质网的Ca2 调节。
①早发型AD 21号染色体基因座与APP基因:St.George-Hyslop等(1987)对8代中分别有48例和54例,6代中有20例和23例的4个常染色体显性遗传高发FAD家系,用2l号染色体DNA探针进行连锁分析。发现早发型AD基因座位于21号染色体21q11.2-21q22.2区。Schellenberg(1991)对21号染色体长臂各区测定,优势对数值(lod score,log of the odd of linkage)仅在一些早发型AD,而D21S13,D21S16和D21S110呈小的正值,其余均呈现负值。提示早发型AD的部分亚群与2l号染色体连锁,而一部分定位尚不完全清楚。
SP的βA4由39~43个氨基酸组成,由APP基因编码,连锁分析该基因也定位于21q11.2-21q22.2区,提示APP基因可能与AD发病有关。APP基因含18个外显子,在转录过程中,因7、8和15外显子选择性剪接,产生相应编码为770、751和695个氨基酸的转录子APP770、APP751和APP695。编码β淀粉样蛋白的是APP基因的16和17外显子。
突变系指基因序列上的突变,结构的改变导致功能的异常,影响其表达的蛋白质的质和量。突变有的是基因部分缺失或全部缺失,有的则是由于外源DNA序列插入,插入发生在外显子上,遗传密码发生改变,结果引起蛋白序列较大变动。最早发现的APP基因突变发生在17外显子的第693密码子,相应于β淀粉样蛋白第22个氨基酸,由谷氨酸变为谷氨酰胺。以后又在第2个早发型AD家系发现717号密码子由缬氨酸变为甘氨酸的突变。
上述APP基因突变均位于βA4区或邻近区域,提示该基因突变可能与βA4形成和AD发病有关。然而21号染色体APP基因突变,仅见于2%~3%少数FAD病人,多数FAD和晚发AD未见此种突变。
②14号染色体基因座与AD3、S182基因:自从1991年发现某些早发型AD家系APP基因突变后,科学家一直致力寻找AD其他相关基因。用短串联重复多态性标记连锁分析,发现某些早发型AD的另一致病基因座位于14号染色体长臂中间14q24.3区。9个家系得到的连锁优势对数值Z=9.01(Z>3有连锁性)。迄今已在11个家系发现早发型AD与14号染色体连锁。有关基因(AD3和S182,不同的研究者对同一基因可能使用不同名称)已被克隆,是一种常染色体显性基因,其突变可能是70%~80%早发型AD的病因。
③1号染色体基因座与STM2或E5-1基因:最近对4个VG(Valga German)家系AD患者进行了1号染色体STM2基因全部编码区的序列分析,结果在141号密码子发现突变,核苷酸由A→T(AAC→ATC)即N141 I突变。这个基因称为STM2或E5-1基因。STM2/E5-1和位于第14号染色体上的S182基因都编码跨膜蛋白,都是常染色体显性遗传。在分子结构,突变发生位点上都有很高的同源性,可能属于一个基因家族,因而将14号染色体AD基因称为早老素-1(Presenilin-2,PS-2)基因S182。70%~80%早发型AD由该基因突变引起。1号染色体上AD基因称为早老素2(Presenilin-2,PS-2)基因,STM2/E5-1与早发型AD的一个小亚组VG有关。早老素的发现是AD研究领域近年来重大突破,已成为AD研究的热点。
(2)晚发型AD:19号染色体基因座和载脂蛋白(Apo E)基因,在更常见的散发性晚发型AD中,Apo E基因(Apo Eε4)是当前分子生物学和分子遗传学研究的热点。已知Apo E是一种血脂运转有关的血浆蛋白,是高血脂和动脉硬化危险因子。脑中Apo E参与脂代谢,在外周神经和中枢神经损伤的再生中可能起部分作用。
在晚发型AD的遗传研究中,发现Apo E基因是晚发型AD的重要危险基因。Apo E基因定位于19号染色体的19q13.2位点,编码的Apo E是一种与脂质转运有关的蛋白质,相对分子质量为34×103,由299个氨基酸组成,富含精氨酸,是低密度脂蛋白颗粒的组成成分之一。在大脑中,Apo E是由星形细胞产生,在脑组织局部脂质的转运中起重要作用,特别是与神经元损伤和变性后,髓鞘磷脂的代谢和修复密切相关。Apo E有3种常见亚型,即E2、E3和E4,分别由3种复等位基因ε2、ε3和ε4编码。Apo E 3种蛋白亚型的差别仅在112和158两个位置的氨基酸不同。这种差异是由于Apo E基因在这两个密码子位置的单碱多态性所致。虽然不同人群中,Apo E各等位基因的分布频率有差异,但ε3最常见,其次是ε4,ε2较少见。正常白人中Apo E等位基因的频率分布为:ε2,8%;ε3,78%;ε4,149%。据报道,正常汉族人群中的分布为:ε2,4%;ε3,83%;ε4,13%。大量研究证实,Apo Eε4等位基因的频率,在家族性和散发性AD中显著升高。家族性AD时Apo Eε4等位基因的频率最高,约为50%,经尸解确诊AD患者的Apo Eε4也比较高,散发性AD的频率为24%~40%。进一步的研究表明,Apo Eε4等位基因与晚发型AD明显关联。携带Apo Eε4等位基因使用AD的风险增加,而且使发病年龄提前。有趣的是Apo Eε2等位基因似乎具有保护作用,携带此基因可减少患病风险,使发病年龄延迟。Apo E等位基因型为ε4/ε4的患病风险最高,可增加4~8倍,ε2/ε3型个体的患病风险小于ε3/ε3个体,ε2/ε4与ε3/ε3型个体的患病风险无明显差异。ε4等位基因只是一个危险因素,携带ε4等位基因并不意味着必定导致AD,有许多AD患者没有ε4等位基因,另外,有不少ε4/ε4基因型的个体即使达高龄也没有痴呆的证据。因此,可能有多种基因或其他因素促使AD发病。各国人群Apo Eε4等位基因频率分布见表3和表4。
最近发现,α-1抗糜蛋白酶(ACT)基因能影响与Apo E基因有关的患病风险。Apo E基因型为ε4/ε4,同时ACT基因型为A/A个体的患病风险显著增加。先前也曾发现Apo E基因型可影响具APP突变的AD患者发病年龄,携ε4/ε4基因型的发病年龄显著提前。Apo E是如何影响AD发病的机制还不能肯定。在实验室,Apo E与tau蛋白和β淀粉样蛋白都能结合。曾经假设Apo E与β淀粉样蛋白结合后能促进老年斑的形成,并影响β淀粉样蛋白的代谢,而Apo EⅡ可防止tau蛋白的过度磷酸化,以避免NFT的形成。到目前为止,已发现多个基因与AD的发病有关。AD病因的这种遗传异质性表明,AD可能并非一种疾病,或许是多种疾病的集合。与晚发型AD患病有关的所谓危险基因还有许多,例如,ACT基因、血管紧张素转换酶基因(ACE)、α2-巨球蛋白基因(a2M)、5-HT转运体基因、线粒体基因等,这些基因的作用尚未阐明。
tau蛋白基因突变可引起家族性额颞叶型痴呆,但到目前为止,还没有发现AD有tau蛋白基因突变。Creutzfeldt-Jacob disease(CJD)或称人海绵样脑病,是一种罕见的痴呆。CJD的年发病率约为百万分之一,可分为遗传性和获得性两型,遗传性CJD占5%~10%。70%的遗传性CJD是由PRNP基因的100 K点突变所致。

以上显示尼日利亚Apo Eε4频率最高,其次为苏丹,我国人群(北京)ε4频率最低。

大约近2/3的AD病人至少有1个Apo Eε4等位基因,遗传上有2个Apo Eε4等位基因的病人,发生AD的可能性是遗传有2个Apo Eε3或Apo Eε2等位基因的8倍。估计全球1/4的人群为杂合子Apo Eε4等位基因。另2%~3%人群为纯合子,纯合子人群发生AD的危险高达90%。Apo Fε4基因不仅有助预测谁可能得AD,而且还存在剂量效应,表现为对发病年龄的影响。Corder等(1994)报道一组迟发型AD家系,每增加1个ε4基因拷贝。AD发病年龄提前7~9年。无ε4基因者平均发病年龄为84.3岁,1个E4基因者为75.5岁,2个者发病年龄68.4岁。
有人对患多种内科病的1000名高龄老人(平均年龄85岁)进行Apo Eε基因检测,发现Apo Eε4基因频率与MMSE认知测验分值下降显著相关。认知测验减分的相对危险性明显受Apo Eε4基因剂量影响。虽Apo E是高脂血症和动脉硬化的危险因素,但Apo Eε4基因剂量并不增加脑血管病相对危险度,然而有Apo Eε4等位基因者脑血管病并发痴呆的危险性上升。该作者还观察了Aoo Eε基因对某些病人认知功能减退的影响,发现有Apo Eε4等位基因者,在6个月随访中,MMSE分值比无Apo Eε4者低得多。
然而,某些含Apo Eε4基因的百岁老人并无痴呆,而且约40% AD病人并没有ε4等位基因。另外对含Apo Eε4基因者长达20年监测未出现AD,因此就目前资料来看,Apo Eε4基因可能只是AD的危险因素而并非AD的病因。像冠心病和肿瘤等多基因疾病一样,环境因素在基因激活或突变中可能起扳机作用。
总之,现有资料表明早发型AD基因座分别位于21、14和1号染色体,相应的基因为APP(与βA4形成有关)、S182(早老素-1,70%~80%早发型AD由该基因突变引起)和STM2/E5-1(早老素-2与VG早发型AD有关)。迟发型AD基因座位于19号染色体,相应的基因为Apo E。综上可见AD具有遗传异质性,表明AD不可能是单基因遗传,而可能是遗传与环境因素相互作用,是一个多基因障碍。遗传传递不可能解释普通人群中多数AD病例。
2.细胞骨架和突触 老年期痴呆的神经元细胞和神经胶质细胞都有结构改变。这些改变包括神经细胞死亡、树突收缩和膨胀、突触减少和神经胶质细胞反应。控制细胞生长和活动的细胞信号通路也会发生变化。细胞骨架包含许多不同大小的聚合体和蛋白质成分。聚合体分为微丝(直径6nm),由肌动蛋白构成;中丝(直径10~15nm),由一种或多种特殊的中丝蛋白构成,不同细胞中各不相同(如神经元中的神经丝蛋白和星形胶质细胞中的胶质纤维酸蛋白);微管(直径25nm),由微管蛋白构成。除形成细胞骨架的丝蛋白外,细胞还有一些与细胞骨架结合的蛋白,这些蛋白能调节神经丝蛋白的聚合和解聚合反应。神经元表达几个与微管结合的蛋白质(MAP),它们分布在复杂的细胞结构中不同部位,例如,MAP-2存在于树突中而轴突中没有,tau蛋白存在于轴突中而树突中没有。
正常老化过程中,细胞骨架中的蛋白质含量没有很大的变化(如微管、肌动蛋白和神经丝蛋白),但细胞骨架排列和细胞骨架蛋白翻译后的修饰有所变化,例如,有些脑区tau蛋白的磷酸化水平增高,特别是与学习和记忆有关的区域(如海马和前脑基底)。另外,有些神经元中由钙调节的细胞骨架蛋白如MAP-2和血影蛋白(spectrin)的水解增多,神经胶质的纤维酸性蛋白增加。对啮齿类动物和人类的不同突触进行多种类型的分析发现,在老化过程中某些脑区的突触数量会减少,但突触体积增大可以弥补这种减少,突触可有“重建”。
AD的重要神经病理变化NFT是皮质和边缘系统神经元内的不溶性蛋白质沉积。在电子显微镜下,构成缠结的蛋白质为双股螺旋丝(PHF)。PHF横向联结紧密、高度不溶。双股螺旋丝的主要成分是高度磷酸化的tau蛋白。tau蛋白的相对分子质量为5万~6万,是一种微管结合蛋白。编码该蛋白的基因位于17号染色体的长臂。tau蛋白对维持神经元轴突中微管的稳定起重要作用,而微管与神经原中的物质转运有关。tau蛋白的氨基酸序列的重要特征是C末端3个或4个重复序列,这些系列组成微管结合位点。tau蛋白过度磷酸化后,其与微管的结合功能受到影响,参与形成NFT、神经元网线(neuronal thread)和老年斑。现在还不清楚tau蛋白是如何磷酸化及如何横向紧密结合以致高度不溶的。蛋白激酶C和谷氨酸能神经元的活性异常可能与tau蛋白的过度磷酸化有关。
NFT是tau蛋白在退化的神经元胞质中形成的细丝状累积。有缠结的神经元微管数量减少,细胞中常有微管结合蛋白如MAP-2和tau蛋白的构型改变。在NFT中tau蛋白被过度磷酸化,可能是由于磷酸化酶活性改变引起去磷酸化减少或脂质的直接氧化和过氧化产物如4-hydroxynonenal的共价键修饰所致。
突触减少是AD的特征之一。突触减少是否发生在细胞死亡前尚无定论,但肯定与神经元退化和死亡有关。越来越多的证据表明,突触内的兴奋性中毒会导致突触退化,这可能会触发神经元细胞的死亡进程。在脊髓树突突触后膜谷氨酸受体聚积明显增加,表明在正常的生理性突触传递中有大量的Ca2 内流。与衰老相关的生化改变(如神经元能量不足和氧化水平增高)和与疾病有关的生化改变(如AD的淀粉样蛋白)会使突触容易受兴奋性中毒损害。
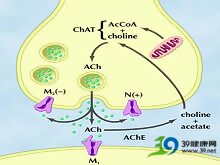
3.中枢神经递质 为探索AD的发病机制,对AD病人中枢神经递质进行了深入研究。Ach(乙酰胆碱)在脑和脊髓的某些神经元中起神经递质的作用。基底前脑神经元的胆碱能神经通路与广泛的皮质区域和海马联结,这些胆碱能神经元与学习和记忆密切相关。20世纪90年代以来,AD脑中神经递质的浓度、受体数量和神经递质合成酶成为AD研究的重点。已明确有很多重要神经递质或调质广泛脱失,特别是在大脑皮质和海马联合区。一般认为,AD的核心症状记忆丧失是由于乙酰胆碱缺失引起,此即AD胆碱功能低下假说。根据为:①AD病人存在广泛nbM胆碱能神经元变性和脱失;②数百例确诊AD病人中,脑中乙酰胆碱及其合成酶胆碱乙酰转移酶(ChAT)水平低下,在一些脑区ChAT可减少到正常水平的30%。ChAT位于胆碱能神经元内,被视为胆碱能神经元的标志,ChAT活性减少,表明胆碱能神经元脱失,虽ChAT活性随增龄而递减,但AD减少最显著。有报道ChAT减少和胆碱能神经元脱失高度相关,也与AD病人生前认知功能和大脑病理改变程度相关,其他递质虽也减少,但未发现相关性;③东莨菪碱和阿托品都是毒蕈碱胆碱能受体拮抗剂,可引起正常青年人记忆和认知功能减退,与AD病人记忆减退相似;④胆碱酯酶抑制剂毒扁豆碱或毒蕈碱受体激动剂槟榔碱(arecoline)能逆转东莨菪碱引起的记忆减退;⑤用兴奋性神经毒素损伤啮齿动物nbM,可引起动物胆碱能功能低下状态,表现为学习和记忆障碍。目前用药物促进神经传递以补偿递质功能不足使帕金森病的治疗有了很大的进展。因此,人们希望发现AD的神经递质缺陷,以便寻找有效的治疗方法。
最早发现的是AD胆碱能神经元系统有特异性的神经递质缺陷。皮质和海马的胆碱乙酰基转移酶(ChAT)减少。ChAT含于前脑底部神经核团的胆碱能神经元突触前襻中。前脑底部神经核团主要由Meynert基底神经核、中隔核组成。这些神经核损害使ChAT减少,以致在皮质和海马合成Ach减少。Ach与近记忆密切相关,而记忆障碍是AD的主要临床表现。有趣的是AD只有投射性胆碱能神经元受损,基底核和脊髓的胆碱能神经元不受影响。人脑皮质中的胆碱能受体主要是M1受体,约占80%。AD的M1受体数基本正常,少数研究发现AD的M2受体减少,而且与胆碱能神经元受损程度相关。M1受体数正常提示AD的胆碱能递质缺陷可能通过代偿机制而调整其功能。有人认为受体数的研究结果可能并不可靠,因为即使受体正常,如果细胞内的第二信使不能激活G蛋白,信息就无法传至细胞内,也就不能有效地激活神经元。
AD患者的胆碱能递质系统损害非常明显,胆碱的运输、Ach合成和释放减少,胆碱能M受体与GTP效应蛋白的结合减少,尤其明显的是M受体激动剂激活皮质神经元细胞膜内的GrP结合蛋白能力下降。神经元中膜脂质过氧化水平增高可导致胆碱能信号受损,因为把培养的皮质神经元暴露于β淀粉样蛋白、Fe2 和脂质过氧化产物4-hydroxynonenal时,会使胆碱能M受体与GTP结合蛋白Gqll的结合能力受损。
氧化压力会导致多巴胺功能不良,氧自由基在帕金森病的发病中起主要作用,但氧化压力在多巴胺信号改变中所起的作用还不清楚。多巴胺受体与GTP效应蛋白的结合存在与年龄相关的损害。大部分AD患者的黑质纹状体系统无明显的多巴胺能神经元脱失。少数研究发现AD患者的脑脊液中多巴胺的代谢产物降低。
去甲肾上腺素和5-HT是脑中主要的单胺能神经递质。去甲肾上腺素神经元主要位于蓝斑,5-HT神经元位于中缝核。这两种神经元都广泛地投射到脑皮质区域。去甲肾上腺素的受体有几种亚型,每一种都能与GTP结合蛋白结合。5-HT受体也有几种亚型,其中有些与GTP结合,有些构成配体性离子通道。AD患者脑中去甲肾上腺素总量和再摄取量都有减少,合成去甲肾上腺素的TH减少,脑干的蓝斑中神经元有脱失。有趣的是蓝斑神经元的脱失仅限于投射至前脑的神经元,而投射至小脑和脊髓的神经元正常,也没有AD的病理改变。蓝斑神经元受损的程度及去甲肾上腺素减少的程度与认知功能减退的程度无关,但与AD的情感症状有关。AD患者缝际核中的神经元有脱失,皮质和脑脊液中5-HT及其代谢的产物浓度有降低。5-HT的改变可能与AD的非认知性精神症状,如抑郁、攻击行为等有关。
人脑中谷氨酸是主要的兴奋性神经递质。谷氨酸激活亲离子受体,引起Ca2 和Na 内流。亲离子的谷氨酸受体过度激活在卒中、AD、帕金森病和亨廷顿病发病机制中起重要作用。随着年龄增大,谷氨酸亲离子受体的水平下降,可能是由于有关的神经元发生退化而引起。谷氨酸能神经传递在神经元死亡、与年龄相关和与疾病相关的脑功能缺损中所起的作用还不大清楚。人脑中主要的抑制性神经递质是γ氨基丁酸(GABA)。在AD等神经退化性疾病中,谷氨酸脱羧酶水平下降,GABA结合位点减少。GABA系统在各种痴呆中作用机制目前还知之甚少。
4.血管病变 与其他器官一样,脑部的血管很容易出现动脉粥样硬化,使血管很容易出现闭塞和破裂,导致卒中。这是血管性痴呆(VD)的最主要的发病机制。脑血管病的最常见后果是神经细胞缺失和认知功能损害。脑灌注下降在认知功能衰退中扮演了重要角色。老年期痴呆的主要原因中,VD居第2位。随着年龄增大,脑血流下降,同时氧和葡萄糖的代谢率下降。脑老化的微血管结构性改变包括管壁基膜和内膜的卷曲增厚等。血管变化的生化因素可能包括氧化后的低密度脂蛋白(low density lipoprotein,LDL)对内皮细胞的损害和炎症反应。内皮细胞的受损使其功能如葡萄糖的转运受影响,炎症反应使巨噬细胞容易通过血脑屏障。当年龄增大时血脑屏障的功能有削弱,包括毛细血管壁变薄和内皮细胞中线粒体数量减少,内皮细胞重要的转运功能受到损害。AD患者大脑的这种损害更为严重。血管破坏,特别是脑微血管的破坏增多,会导致了神经元损害和认知功能损害。载脂蛋白E(Apo E)的多态性与AD和动脉硬化的危险性增高都有联系,携ε4等位基因者患这两种疾病的危险性都增高,而携有ε2等位基因的则危险性下降。这种联系表明,血管病变改变在AD的神经退化过程中起重要作用。
5.淀粉样蛋白 AD的病理特征老年斑为神经元炎症后的球形缠结,其中包含退化的轴突和树突,伴有星形细胞和小胶质增生。此外,还含有多种蛋白酶。老年斑的中心是β淀粉样蛋白,它是β淀粉样前体蛋白(APP)的一个片段,含39~43个氨基酸。在许多脑区可见不成熟的老年斑,不成熟的老年斑只有β淀粉样蛋白,而没有神经炎性反应。APP是一种跨膜蛋白,尾部位于细胞内,其余大部分位于细胞外。正常的APP代谢是在靠近细胞膜处的α位点被α分泌酶(α-secrtetase)切断,酶切位点正好在β淀粉样蛋白片段的中央,故正常的代谢途径不产生β淀粉样蛋白,只产生一条比较长的APP片段,称为可溶性APP(sAPP),此物质具有细胞营养作用。另外还有2个酶切位点,即β分泌酶(β-secretase)和γ分泌酶(γ-secretase)位点。β位点位于a位点的外侧,γ位点位于α位点内侧的细胞膜中。这两个位点酶切后形成39~43个氨基酸的β淀粉样蛋白。然而,现在还不能以上述机制解释全部AD病例,APP代谢为何会转为产生β淀粉样蛋白的可能机制有3个:①某些家族性AD由于APP基因突变致使APP蛋白结构改变;②APP基因正常时,可能与多种因素有关,例如含β淀粉样蛋白的APP片段可见于溶酶中,故淀粉样蛋白可能经溶酶体中的肽酶和蛋白酶的剪切而分泌出来;③多种损伤的结果,多种实验性损伤如前脑底部去皮质神经元支配的损伤可使老鼠的APP合成增加。人类严重脑外伤可见弥漫性老年斑。神经元内氧化压力增加或能量代谢削弱会使APP水解过程改变,促进β淀粉样蛋白和纤维形成。AD患者脑中β淀粉样蛋白的积聚与神经元细胞的衰亡和认知功能损害的严重程度大致平行。
β淀粉样蛋白沉积对神经元有毒性作用,通过自由基、刺激细胞死亡程序或刺激胶质细胞产生肿瘤坏死因子等毒性物质而使神经元死亡。细胞培养表明β淀粉样蛋白具有神经毒性,能使神经元易于受代谢、兴奋性物质和氧化剂破坏。β淀粉样蛋白形成纤维的倾向性与其神经毒性作用有关。防止纤维形成的物质,如刚果红色染料能阻断β淀粉样蛋白的毒性作用。β淀粉样蛋白引起的神经元损害包括胞膜中的脂质发生过氧化,结果产生一种有毒的乙醛叫4-hydroxynonenal。4-hydroxynonenal能对某些膜蛋白进行共价修饰,而这些蛋白与保持胞膜及其能量代谢如Na -K 泵、Ca2 -ATP酶、葡萄糖转运体和谷氨酸转运体的动态平衡有关。神经元中β淀粉样蛋白引起的氧化压力不仅会导致细胞内Ca2 浓度增高、线粒体功能障碍和细胞凋亡(一种细胞死亡的形式,特征是细胞皱缩,细胞核浓缩,DNA片段形成),还会导致细胞骨架发生变化。这些变化与NFT中所见到的类似。抗氧化剂如维生素E和谷胱甘肽能防止细胞凋亡和NFT。某些神经营养因子(脑内产生的细胞信号蛋白)通过诱导抗氧化酶基因的表达来保护细胞不受β淀粉样蛋白的损害。除了参与神经退化过程,β淀粉样蛋白还会损害神经递质的信号通路。β淀粉样蛋白不仅会损害乙酰胆碱M型受体与三磷酸鸟嘌呤效应蛋白的结合,还会抑制乙酰胆碱的产生。β淀粉样蛋白可损害脑血管,并且造成脑实质中营养运输不足。AD患者的脑血管中有大量的β淀粉样蛋白沉积。β淀粉样蛋白还会损害内皮细胞的葡萄糖运输,使这些细胞的屏障作用下降。β淀粉样蛋白在内皮细胞中诱导氧化压力,造成细胞退化和死亡,同时也会促进脑内的炎症过程。
由于转基因技术的发展,目前能用转基因技术研究基因产物过量表达或基因突变的作用。现在已能通过动物模型展示某些类型AD的病理变化。通过转基因技术将家族性AD的APP变异基因植入老鼠,其人脑能产生β淀粉样蛋白沉积、神经炎性老年斑、突触减少、星形细胞和小胶质增生、神经元退化及神经元程序性坏死。
有学者用β淀粉样蛋白的抗体作ELISA检测,从AD患者的脑脊液中测出多种异型肽片段,包括27~43个氨基酸残基的片段,都以天冬氨酸为起始端。Nakamura等发现早发型AD(平均59岁)患者脑脊液中β淀粉样蛋白水平显著增高,而晚发型AD与正常对照组无显著差异,认为β淀粉样蛋白与早发型AD相关。Motter等以β淀粉样蛋白42为同定指标,检测37例AD的脑脊液,结果β淀粉样蛋白42含量显著低于对照组及其他神经系统疾病患者,但tau蛋白显著增高,淀粉样蛋白总量无显著差异。
有关β淀粉样蛋白的研究,近年来最突出的进展有两个,其一是已分离出β和γ分泌酶,为开发有可能抑制该酶和老年斑形成的药物提供了化学基础。这两种分泌酶的抑制剂正在开发研究中,今后有可能用β和γ分泌酶抑制剂来阻止老年斑的形成。其二是用来对抗老年斑的β淀粉样蛋白“疫苗”已在动物实验获得成功,用β淀粉样蛋白“疫苗”接种APP转基因老鼠,可防止或减少老年斑的形成。目前,β淀粉样蛋白“疫苗”已进入临床实验阶段。非常遗憾的是接种β淀粉样蛋白“疫苗”的志愿者中有少数出现了“脑炎样反应”,实验不得不暂时停止,以便分析原因。可以相信,如果这两种新疗法获得成功,将使AD的预防和治疗效果大为改观。
6.脂质、核酸和自由基 某些痴呆患者特别是AD患者脑中不溶性蛋白质积聚增多,细胞骨架tau蛋白和β淀粉样蛋白是与此关系最为密切的两种物质。神经元细胞中脂质含量比较高,尤其是在有髓胶质细胞如少突胶质细胞中,所以大致有一半的脑净重量是由脂质构成。AD患者脑中的髓壳脂质减少,包括脑苷磷脂、磷酰胆碱、磷酸乙醇胺和鞘磷脂逐渐减少,而细胞内破坏性脂质增加。破坏性细胞膜脂质的积累表现为神经元中脂褐素颗粒增多。
AD患者脑中DNA和RNA的含量减少。虽然在Meynert基底神经核、大脑皮质的几个区域和有些脑神经核内的RNA水平下降,但在海马下脚区RNA水平升高。蛋白质的氧化水平通常用蛋白质碳酰基来测量。脑组织研究发现AD和帕金森病患者的易感脑区,特别是退化的神经元中蛋白质氧化水平升高,碳酰基显著升高。蛋白质糖残基增多称为糖化,蛋白质糖化会增加细胞的氧化压力。老年斑和NFT的主要成分中的β淀粉样蛋白和tau蛋白就是两种过度糖化的蛋白质。
在神经退化性疾病中,线粒体DNA会出现进展性和广泛性的氧化。HD患者的纹状体和AD患者的海马和易感皮质区DNA的损害已得到证明,特别是8-羟基鸟嘌呤的水平升高。这种DNA损害可能是由带有羟氢氧自由基和过硝酸盐根的几个反应性氧分子造成。线粒体DNA损害的原因是大部分自由基在线粒体产生。细胞培养试验发现,线粒体DNA受损会导致线粒体中电子传输失败并减少ATP的生成,还会导致线粒体的钙屏蔽功能受损,增加神经元对兴奋性毒性物质和代谢性伤害的敏感性。
自由基是指原子核外层轨道带有不对称电子的分子。在生物体内,各种携氧分子是自由基的主要品种。细胞中氧自由基主要来源于线粒体,这是因为线粒体内的电子传递过程中可产生超氧阴离子自由基(O2-)。超氧化歧化酶(MnSODt和Cu-Zn-SOD)把O2-转化为过氧化氢(H2O),H20然后在Fe2 和Cu2 的催化下,通过Fenton反应转变为O2-H。过氧化亚硝酸盐(peroxynitrite,0N00-)来自氧化亚氮(NO2-)和O2-的互相作用。刺激NO生成的主要因素是Ca2 水平增高,它与钙调蛋白结合并激活NO合酶。虽然O2-H和ONOO-能对蛋白质和DNA造成直接损害,但它们破坏细胞的主要形式是攻击细胞膜上的脂肪酸,并发动脂质过氧化过程。
来源于线粒体的氧自由基在细胞的逐渐氧化损害起主要作用。脑供能和代谢损害可导致氧自由基的产生增多。限制热量摄入似乎可以减少线粒体氧自由基的产生。用限制热量的饮食饲养大鼠(热量减少30%~40%),发现许多非神经性组织中细胞氧化压力降低(如蛋白质、脂质和DNA的氧化减少)。最近的研究发现,用限制热量饮食饲养的啮齿类动物脑中,氧化压力也减少。在有神经退化性疾病,除了氧自由基的损害,一些引起氧化压力的因素也起重要作用,例如,AD患者β淀粉样蛋白的不断产生和积聚是造成神经元氧化压力增高的关键因素。细胞培养证实,β淀粉样蛋白可引起神经元膜脂质过氧化,这些氧化压力会破坏神经元,导致神经元死亡。
7.线粒体与能量代谢 痴呆的神经元异常代谢增多,在神经退化性疾病所致痴呆中更为严重。在AD患者脑细胞中,几种能量代谢过程中酶的活性严重减少。其中与线粒体氧化代谢有关的3种酶是丙酮酸脱氢酶复合物、α-酮酸脱氢酶复合物和细胞色素C氧化物。这些酶的活性严重不足可能是由于编码这些酶的DNA受到了氧化性损害。另一导致神经元能量代谢下降的因素是神经元细胞膜上转运葡萄糖的蛋白质功能受损。对AD患者死后脑组织的研究表明葡萄糖转运体数量减少,对海马神经元和突触体的研究发现与AD病理过程有关的损害(β淀粉样蛋白和氧自由基)会削弱葡萄糖的转运。葡萄糖运输减弱和线粒体功能不良导致ATP损耗,致使神经元易受兴奋性毒性物质损害。
线粒体功能障碍与多种神经退化性疾病有关。在帕金森病中有明显的线粒体酶功能不良,包括复合物Ⅰ和α酮酸脱氢酶复合物活性下降。把培养中的细胞暴露于与帕金森病发病有关的伤害因素,如N-甲基4-苯基四氢吡啶(MPTP)和铁时会引起线粒体功能障碍。AD的易损脑区中细胞色素C氧化酶和α-酮酸脱氧酶的活动水平显著减少。有趣的是在AD患者中,也能见到非神经元细胞如血小板和纤维原细胞有线粒体损害。近年来的研究发现,某些AD病例的线粒体DNA有遗传性改变,这些改变是母体遗传,因为在受精过程中,没有线粒体从精子传递给卵子。把AD患者血小板中的线粒体引入培养中的神经瘤细胞时,氧化压力增高,这表明线粒体在改变AD脑神经元氧化压力方面起重要作用。把培养中的海马神经元或皮质突触体暴露于与AD发病有关的损害因素时(如β淀粉样蛋白和4-hydroxynoncnal),会导致线粒体功能障碍。
8.神经元离子内稳态 神经元的兴奋性是与许多其他类型细胞不同的特征,兴奋性由复杂的神经递质传递和离子通道构成。神经元有电压依赖性的钠通道,还有多种类型的钙和钾通道,在不同的神经元和不同的细胞间隔之间有不尽相同离子通道。例如,胞体上为L型Ca2 通道,树突上为N型Ca2 通道,突触前膜为T型Ca2 通道。据研究,细胞内钙离子保持在10-8mol范围,短期内细胞内钙离子浓度较大改变(如缺血)或长时间小的改变(如老年痴呆),慢性钙离子超载,都可以引起AD神经细胞变性和死亡。另外,神经元还有膜离子“泵”,它在神经元受刺激后恢复离子平衡的过程中起重要作用。实验研究发现,痴呆的神经元有许多电生理参数的改变,例如,引起动作电位的阈电位水平增高,海马神经元中后超级化电位增高,海马中突触传递的受损等。这些改变与相应的神经环路的功能降低有关,例如,听神经功能下降导致听觉受损,海马环路功能下降导致认知功能受损。
在神经系统中,Ca2 在调节神经元的存活和可塑性方面起根本性作用,Ca2 不仅调节神经递质和神经营养因子对神经生长、突触生成的作用,它还调节神经系统细胞的生存。在成熟的神经系统,前突触Ca2 内流刺激神经递质释放,后突触Ca2 的内流与学习和记忆过程密切相关。老年人的血浆Ca2 、ATP酶活性下降和Ca2 结合蛋白水平降低,相反,AD患者的神经元中,钙依赖蛋白酶的活性增高。在衰老和神经退化性疾病中,削弱神经元中Ca2 动态平衡的因素可能有氧化压力和代谢损害。的确,动物试验和细胞培养研究发现,氧化(如Fe2 、β淀粉样蛋白)和代谢损害(如葡萄糖缺乏、线粒体毒素)可破坏神经元Ca2 的动态平衡。
9.神经内分泌 神经内分泌的变化对脑老化和神经退化性疾病有重要作用,类固醇激素、糖皮质激素和雌激素的分泌变化尤为重要。AD患者的下丘脑-垂体-肾上腺素轴的调节会发生改变,使血浆糖皮质激素水平增高。生理或心理应激会造成糖皮质激素水平增高,导致海马神经元对缺血和兴奋性伤害更为敏感,甚至会引起神经元的死亡。这些发现表明糖皮质激素对急性神经疾病(如卒中)和慢性退化性疾病(如AD)有负性的作用。雌激素对脑老化有保护作用。流行病学研究发现,绝经后妇女接受雌激素替代治疗可显著降低患AD的危险性。另外服用雌激素的妇女认知功能也更好。动物和细胞培养研究表明,17β-雌二醇能保护神经元免受缺血、缺氧或与AD有关因素的破坏。
10.神经营养因子 神经科学研究发现中枢存在神经营养因子。它是天然存在的物质,神经系统细胞会产生一些蛋白质,它们能促进细胞存活和生长,并能保护神经元避免伤害和死亡,这些蛋白质就是神经营养因子。神经营养因子主要有神经生长因子(NGF)、纤维原细胞生长因子(bFGF)、脑源性神经营养因子(BDNF)和类胰岛素生长因子(IGF)。神经营养因子在细胞培养中能保护神经元,在活体中能对抗与神经退化性疾病有关的一些损害。据报道AD病人含量减少,从而导致神经细胞死亡和功能障碍。例如,在细胞培养中bFGF能保护海马和皮质神经元不受代谢、氧化和兴奋性因素的损害。在动物卒中模型中,bFGF能减少脑坏死。BDNF能保护多巴胺神经元不受MPTP(PD动物模型)的毒性伤害,bFGF和sAPP能保护海马神经元不受β淀粉样蛋白的毒性伤害(AD动物模型)。神经营养因子的作用有两种机制,即增加细胞对氧化压力的耐受性和稳定细胞Ca2 的动态平衡。神经营养因子的这些作用似乎是通过诱导抗氧化酶如超氧化歧化酶和谷胱甘肽过氧化酶和钙调蛋白表达的结果。
神经营养因子一个有趣的特性是它们表达随着神经环路活动的增加而增加。细胞培养和活体研究表明,这种活动依赖性的神经营养因子在促进神经元存活和神经元突的生成中起重要作用。在富于挑战的环境中喂养的老鼠,其海马和某些脑皮质区神经元的树突和突触数量增多。流行病学资料提示,积极脑力活动的人患AD的危险性下降,这表明大脑存在“用而不废”的情况。经常的脑力活动可促进神经营养因子的合成,进而促进神经元的生长和可塑性。
11.其他
(1)微量元素:早期研究发现铝及硅在NFT和SP含量较正常老人高,长期以来一些科学家致力于铝对本病的可能作用,流行病学研究有报道水中含大量铝的地区其居民患本病的危险性增高。一般认为高浓度铝及硅在脑中堆积是细胞死亡的结果而不是原因。最近Garruto等(1994)提出肾透析病人发生痴呆的危险性高,脑铝水平及老年斑都较多。虽无明显NFTs,但确有tau蛋白加工异常和见于AD病人的磷酸化tau蛋白沉着。体外研究也表明铝可能促进tau蛋白磷酸化,导致淀粉样蛋白生成,看来铝在AD发病中的作用尚存有争议。
(2)慢病毒感染:已知与AD类似的一种脑病克-亚综合征(Creutzfeldt-Jacob病)是由慢病毒引起的,有人推测AD也可能如此。但迄今未能在AD脑组织中分离出病毒,也未发现与其他病毒性疾病类似的免疫反应。
(3)脑外伤:主要指伴有意识障碍的脑外伤,拳击家痴呆为脑外伤假说提供了佐证,有报道1059例AD中87例有脑外伤史,比值比OR为1.8(95%可信限,3~2.7),我国1988~1990年对126例符合:ICD-10 AD病人对照研究,危险因素条件Logistic回归分析,脑外伤未能列入危险因素之中。
(4)免疫:人体的免疫系统可以保护机体免遭有害异物侵袭,如果产生针对自身重要细胞的抗体,错误地攻击自身组织,称为自身免疫反应。此现象也可发生于脑部,因此有人提出AD是一种免疫系统疾病。理由为AD家族病人中自身免疫性疾病如甲状腺炎的患病率高;老年斑核心发现有免疫球蛋白,AD患者脑中抗神经抗体滴度较高。
(5)兴奋性毒素:近年对神经细胞死亡机制有更多了解,据研究谷氨酸和天门冬氨酸是中枢兴奋性神经递质,具有重要生理功能。如大量释放可以造成组织损害,被称为兴奋性毒素(excitoxins),它通过作用于氨基酸一个受体亚型N MDA(N-methyl-D-aspartate),引起去极化,Cl-被动流入增多,大量阳离子和水进入细胞,导致神经元溶解及一系列生化改变,包括酯酶、蛋白酶被激活引起神经细胞死亡。
总之,AD病因和发病机制十分复杂,目前尚未阐明。
- 建议就诊科室
- 精神病科
- 可能疾病
- 1、 妄想症,可能伴随迫害妄想、机械地重复周围人的言语或行为、被害妄想等症状,应去精神病科或心理咨询就诊。
- 2、 硬皮症伴发的精神障碍,可能伴随焦虑、神经症性抑郁、被窃或疑病妄想等症状,应去风湿科或心理咨询就诊。
- 3、 偏执性精神病,可能伴随被害妄想、诉讼狂、夸大狂等症状,应去精神病科或心理咨询就诊。
- 4、 急性妄想发作,可能伴随迫害妄想、偏执性妄想、幻觉等症状,应去心理咨询或精神病科就诊。
- 易混淆症状
- 迫害妄想、偏执性妄想、被害妄想
- 相关检查
- 1、MRI,核磁共振成像(MRI)对 神经系统的病变、脊髓脊椎的病变、心脏大血管的病变、腹部盆腔脏器的检查及关节软组织病变有诊断意义。





